
Рис. 4. 1. Пропорция лобных долей по отношению ко всему мозгу у разных видов приматов и обезьян
Мозг состоит из сотен миллиардов клеток (нейронов и глиальных клеток), сложным образом взаимосвязанных проводящими путями (дендритами и аксонами). Существуют различные типы нейронов и глиальных клеток. Некоторые из проводящих путей между нейронами локальны, они связывают непосредственных «соседей». Однако другие являются длинными, соединяющими отдаленные друг от друга нейронные структуры. Эти длинные проводящие пути покрыты белой жировой тканью, миелином, который способствует прохождению электрических сигналов, порождаемых внутри нейронов (потенциалов действия). Нейроны и короткие локальные связи вместе образуют серое вещество, а длинные миелиновые проводящие пути образуют белое вещество. Каждый нейрон взаимосвязан с мириадами других нейронов, в результате чего образуются сложные конфигурации взаимодействия. Таким образом, сеть умопомрачительной сложности сконструирована из относительно простых элементов.
Принцип достижения большой сложности через многократные преобразования простых элементов представляется универсальным, и этот принцип разнообразными путями воплощен в природе (и культуре). Подумайте, например, о языке, где тысячи слов, предложений и повествований сконструированы из нескольких дюжин букв; или подумайте о генетическом коде, где практически бесконечное число вариантов может быть реализовано благодаря комбинированию конечного числа генов.
Хотя сигнал, порождаемый внутри нейрона, является электрическим, коммуникация между нейронами принимает химическую форму. Многочисленные биохимические системы мозга переплетаются друг с другом, образуя структурную сложность, описанную выше. Биохимические субстанции, называемые нейротрансмиттерами и нейромодуляторами, делают возможным взаимодействие между нейронами. Электрический сигнал (потенциал действия) порождается в теле нейрона и перемещается по аксону, пока не достигает терминала, точки контакта с дендритом — проводящим путем, ведущим к другому нейрону. В точке контакта имеется щель, называемая синапсом. Прибытие активного потенциала высвобождает небольшие количества химических субстанций (нейротрансмиттеров), которые перемещаются через синапс как плоты по реке и прикрепляются к рецепторам, высокоспециализированным молекулам на другой стороне щели. Совершив это, нейротрансмиттеры затем распадаются в синапсе с помощью специализированных катализаторов. Между тем, активация постсинаптических рецепторов выражается в другом электрическом явлении, постсинаптическом потенциале. Множество постсинаптических потенциалов, возникающих совместно, имеет результатом другой потенциал действия, и процесс повторяется тысячи и тысячи раз вдоль как параллельных, так и последовательных проводящих путей. Это позволяет кодировать информацию колоссальной сложности.
Мы постоянно получаем информацию о все новых типах нейротрансмиттеров. К сегодняшнему дню открыто несколько их десятков; к их числу например, относятся: глутамат, гамма-аминомасляная кислота (ГАМК), серотонин, ацетилхолин, норадреналин и дофамин. Некоторые нейротрансмиттеры, например глутамат или ГАМК, находятся в мозге практически везде. Другие нейротрансмиттеры, как дофамин, ограничены только определенными частями мозга. Каждый нейротрансмиттер может связываться с различными типами рецепторов, некоторые из которых являются повсеместными, а другие специфичны для соответствующей области мозга.
Мозг может мыслиться как сопряжение двух крайне сложных организаций, структурной и химической. Это сопряжение ведет к экспоненциальному возрастанию общей сложности системы. Она, в свою очередь, еще более повышается повсеместными петлями обратной связи, где активность источника сигнала модифицируется его целью, на локальном и глобальном, структурном и биохимическом уровнях. В результате мозг может производить практически бесконечное множество различных активационных структур, соответствующих практически бесконечным состояниям внешнего мира. Нейрон представляет микроскопическую единицу мозга и формы связи между нейронами представляют микроскопическую организацию мозга.
Когда организм подвергается воздействию новых конфигураций сигналов внешнего мира, сила синаптических контактов (легкость прохождения сигнала между нейронами) и локальных биохимических и электрических свойств постепенно меняются в сложных распределенных комплексах. Это и есть процесс обучения, как мы его представляем сегодня1.
Нейроны группируются в связанные структуры, ядра и области. Каждая структура состоит из миллионов нейронов. Ядра и области представляют макроскопические единицы мозга, и картина связности между ними представляет макроскопическую организацию мозга. Мозг является в высокой степени взаимосвязанной системой, и архитектура главных связей между его ядрами и полями представляет удобную картину всей системы «с высоты птичьего полета».
Для эвристических целей я прибегаю к метафоре дерева. У дерева есть ствол и ветви. Ветви разделяются на ветки. На концах веток находятся плоды. В некотором смысле, мозг организован подобным образом. Можно думать о мозге как о «дереве возбуждения и активации». Его ствол отвечает за общее физиологическое возбуждение и активацию, необходимые для функции различных мозговых структур, плодов. Это анатомическая ось мозга, ствол мозга. Массивное повреждение ствола мозга нарушает сознание и может привести к коме.
Внутри компактного стволового центра мозга содержатся многочисленные ядра, на которых строится сложная система проводящих путей. Во многих случаях ядра и их проекции являются биохимически специфическими, привязанными к определенному нейротрансмиттеру; в других случаях они являются биохимически сложными, включающими различные нейротрансмиттеры. Это ветви и ветки «дерева активации». Каждая ветвь содержит проекционные связи с определенной частью мозга, гарантируя ее активацию. Несколько десятилетий тому назад было обычным описывать эти ветви суммарно как восходящую ретикулярную активизирующую систему (ARAS)2. Сегодня во все возрастающей степени удается идентифицировать ее отдельные нейроанатомические и биохимические компоненты, изучать эти компоненты раздельно. Повреждение любой отдельной ветви не разрушит сознание в глобальном смысле, но будет препятствовать специфической функции мозга. Каждая ветвь дерева возбуждения проецируется на различные компоненты мозга, каждый из которых обладает своим собственным набором функций.
В мозге имеется множество подкорковых структур. В ходе эволюции подкорковые структуры развивались раньше коры головного мозга, и на протяжении миллионов лет они направляли сложное поведение различных организмов. У современных рептилий и даже птиц новая кора представлена лишь минимально3. В филогенетически древнем, «некорковом» мозге могут быть выделены два класса структур: таламус и базальные ганглии. На ранней стадии эволюции центральная нервная система разделилась на две боковые половины. Поэтому каждая из описываемых здесь структур мозга состоит из двух одинаковых половин: левой и правой.
Несмотря на некоторое функциональное пересечение, таламус и базальные ганглии были наделены отчетливо различными функциями. В древнем, предкорковом мозге таламус отвечал большей частью за получение и переработку информации из внешнего мира, а базальные ганглии отвечали за моторное поведение и действие. Таким образом, разделение восприятия и действия в архитектуре мозга представляется с самого начала основополагающим. Дерево активации разделяется на два основных ответвления, одно проецируется отдельно на подкорковые механизмы восприятия (дорзальная ветвь), другое — на подкорковый субстрат действия (вентральная ветвь).
Часто рассматриваемый как единая структура, таламус фактически является совокупностью многих ядер. Некоторые из них отвечают за переработку различных типов сенсорной информации: зрительной, звуковой, тактильной и тому подобное. Другие ядра таламуса отвечают за интегрирование разнообразных типов сенсорной информации. В таламусе представлена комплексная иерархия входной интеграции. Дорзомедиальное таламическое ядро находится на вершине этой иерархии и тесно связано с префронтальной корой. Другие таламические ядра, обнаруживаемые вокруг средней линии, являются неспецифическими, отвечающими за различные формы активации4.
Тесно связана с таламусом структура, называемая гипоталамусом. В то время как таламус отслеживает внешний мир, гипоталамус отслеживает внутренние состояния организма и помогает поддерживать их в рамках адаптивных, гомеостатических параметров. Гипоталамус также является собранием различных ядер, каждое из которых соотносится с различным аспектом гомеостазиса: прием пищи, прием жидкости, температура тела и так далее. Вместе таламус и гипоталамус называются диэнцефалон5.
Базальные ганглии включают хвостатое ядро, подушку зрительного бугра и бледный шар. В предкорковом мозге эти структуры играли центральную роль в инициировании действий и в управлении движениями. В развившемся мозге млекопитающих базальные ганглии находятся под особенно жестким контролем со стороны лобных долей и работают в сотрудничестве с ними. Фактически, сотрудничество настолько тесное, что я склонен думать о хвостатом ядре как о части «больших лобных долей».
Структура, называемая амигдала (миндалина), также рассматривается как одно из базальных ядер, но она обслуживает несколько иную функцию. Амигдала регулирует те взаимодействия организма с внешним миром, которые являются решающими для выживания индивида и вида: решения атаковать или скрыться, спариваться или нет, заглатывать или нет. Она предоставляет быструю, прекогнитивную, аффективную оценку ситуации под углом зрения ее значения для выживания6.
Мозжечок является большой структурой, прикрепленной к задней части (или, как скажет нейроанатом, к дорзальному аспекту) ствола мозга. Его анатомия в миниатюре параллельна анатомии всего мозга: стержень, называемый червем мозжечка, и два мозжечковых полушария. Мозжечок важен для движений, в особенности для координации тонких движений с сенсорной информацией. Но последние исследования показали также, что мозжечок тесно связан с лобной корой и участвует в сложном планировании7.
На относительно поздней стадии эволюции мозга начала развиваться кора головного мозга, сначала археокортекс, затем палеокортекс8. Они включают гиппокамп и цингулярную кору. Гиппокамп, «морской конек», составлен из двух длинных структур, прикрепленных внутри височных долей (или, как сказал бы нейроанатом, к их медиальному аспекту). Гиппокамп играет решающую роль для памяти. Некоторые ученые убеждены в том, что он специально посвящен овладению пространством9. Я полагаю, что это узкий взгляд, подсказываемый экспериментами над животными, где пространственное обучение является единственно возможной парадигмой для изучения памяти. У людей гиппокамп также задействован в других формах памяти, таких как словесная память10.
Цингулярная кора (кора поясной извилины) прикрепляется к внутренней поверхности полушарий, лежащей над мозолистым телом. Ее функция не вполне ясна, но она связана с эмоциями. Совместно с амигдалой и гиппокампом, цингулярная кора входит в так называемую лимбическую систему11. Это несколько устаревшее понятие, предполагающее функциональное единство этих структур, эвристичность которого все больше ставится под сомнение. Передняя цингулярная кора, по-видимому имеющая дело с неопределенностью, тесно связана с префронтальной корой12. В некотором смысле, она также является частью больших лобных долей.
Наконец, на сцену вышла новая кора (неокортекс)13, тонкая мантия, охватывающая мозг, с характерной сморщенной поверхностью, напоминающей грецкий орех. Корковая мантия имеет свою собственную сложную организацию. Она состоит из шести слоев, каждый из которых характеризуется своим собственным нейронным составом. Определенные части неокортекса организованы в вертикальные «колонки», рассекающие эти слои и представляющие отдельные функциональные единицы. Появление новой коры радикально изменило способ переработки информации и наделило мозг значительно большей вычислительной силой и сложностью. Разделение на две латеральные симметричные системы продолжается внутри коры вплоть до двух полушарий мозга. Различие между системами «восприятия» и «действия» также сохраняется на уровне новой коры: задняя часть коры посвящена восприятию, а передняя часть действию. Но несмотря на эти разделения, новая кора значительно более взаимосвязана, чем ее подкорковые предшественники. Как мы увидим позднее, это могло иметь адаптивное обоснование.
Появление новой коры радикально изменило «баланс сил» внутри мозга. Древние подкорковые структуры, которые использовались для выполнения определенных функций независимо друг от друга, теперь оказались в подчинении у неокортекса и приняли на себя подсобные функции в тени нового уровня нейронной организации. Для ученого, пытающегося понять эти функции, это составляет источник неясности. Функции, для выполнения которых эти подкорковые структуры сформировались до появления коры, вероятно, не совпадают с функциями, которые они выполняют сегодня, в полностью кортикализованном мозге. И поэтому, парадоксальным образом, наше понимание корковых функций во многих отношениях более точно, чем наше понимание функций таламуса или базальных ганглиев, несмотря на тот факт, что кора, в некотором смысле, более «продвинута».
Новая кора (неокортекс) состоит из различимых областей, называемых цитоархитектоническими полями, каждое из которых характеризуется своим собственным типом нейронной композиции и формами локальной связности. Новая кора выполняет разнообразные функции, но между ее различными функциями и цитоархитектоническими областями нет простых отношений. Она состоит из четырех основных долей, каждая из которых связана со своим типом информации. Затылочная доля имеет дело со зрительной информацией, височная доля имеет дело со звуками, теменная доля имеет дело с тактильной информацией, и лобная доля имеет дело с движениями.
На очень поздней стадии эволюции коры появились два важных феномена: возникновение языка и быстрое развитие управляющих функций. Как мы увидим, язык находит свое место в новой коре, присоединяясь одновременно к различным областям коры. И управляющие функции возникают как командный пункт мозга в передней части лобных долей, префронтальной коре. Лобные доли претерпевают взрывную экспансию на поздней стадии эволюции.
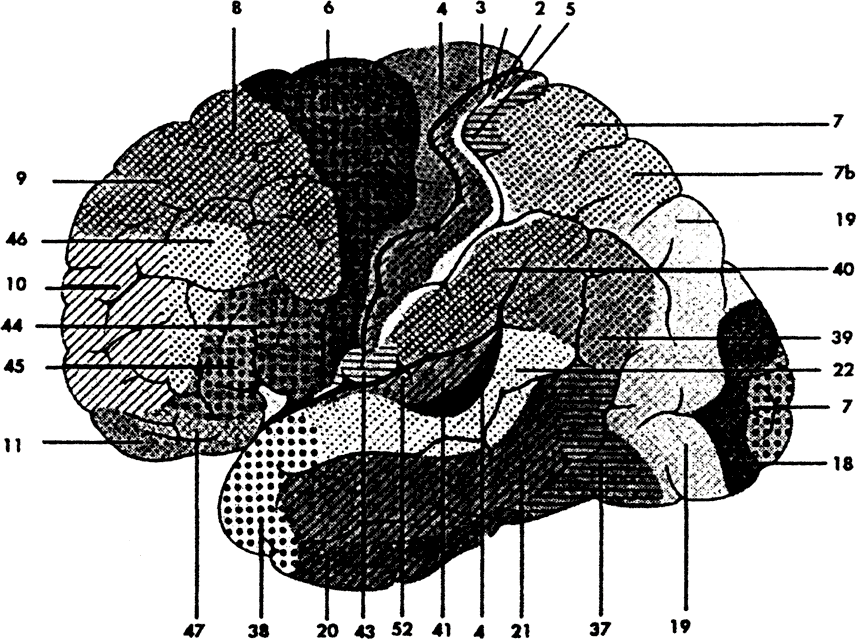
Согласно Корбиниану Бродману14, префронтальная кора или ее аналоги занимают 29% всей коры у людей, 17% — у шимпанзе, 11.5% — у гиббонов и макак, 8.5% — у лемуров, 7% — у собак, и 3.5% — у кошек (рис. 4.1). Существуют различные методы определения префронтальной коры относительно других корковых областей. Один из таких методов базируется на так называемых цитоархитектонических картах, картах коры, составленных из морфологически различных пронумерованных областей мозга (рис. 4.2). Эти кортикальные области называются «зонами Бродмана» — по имени автора наиболее широко используемой цитоархитектонической карты15. Согласно этому определению, префронтальная кора состоит из Бродмановых зон 8, 9, 10, 11, 12, 13, 44, 45, 46 и 4716. Префронтальная кора характеризуется преобладанием так называемых гранулярных нейронных клеток, обнаруженных главным образом в слое IV17.
|
|
Рис. 4. 1. Пропорция лобных долей по отношению ко всему мозгу у разных видов приматов и обезьян |
Альтернативный, но приблизительно эквивалентный метод выделения префронтальной коры основан на ее подкорковых проекциях. Для этой цели обычно используется особая подкорковая структура — дорзомедиальные ядра таламуса, которая в некотором смысле является точкой совмещения, «вершиной» интеграции, возникающей внутри специфических таламических ядер. Тогда префронтальная кора определяется как область, получающая проекции от дорзомедиальных ядер таламуса. Иногда префронтальная кора определяется через ее биохимические проводящие пути. Согласно этому определению, префронтальная кора задается как область, получающая проекции из мезокортикальной дофаминовой системы. Разнообразные методы определения префронтальной коры очерчивают примерно одни и те же территории. Это показано на рисунке 4.3.
 |
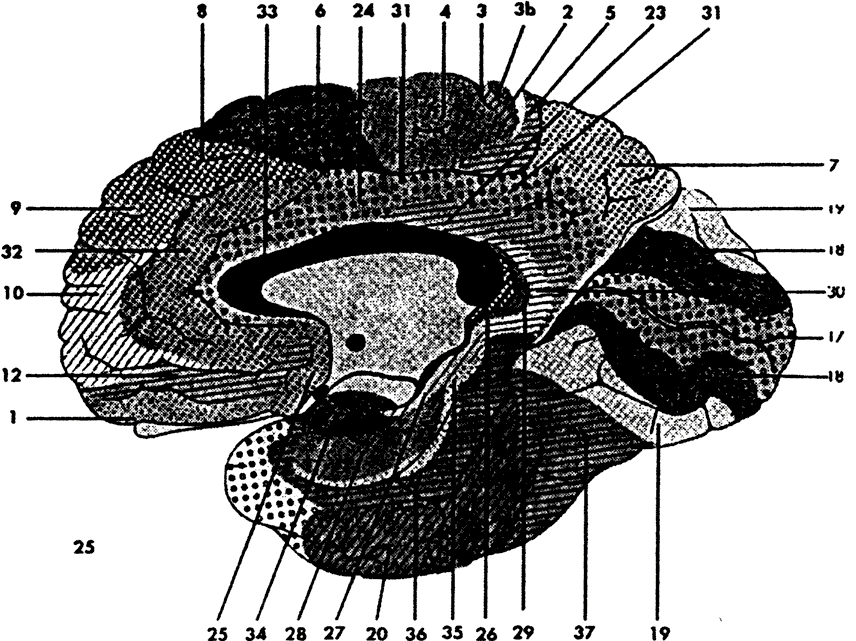 |
Рис. 4.2. Карта архитектонических зон коры по Бродману. (Адаптировано из Roberts, Leigh, Weinberger, 1993.) |
По любопытной параллели между эволюцией мозга и эволюцией науки о мозге (мы еще не один раз вернемся к этому), интерес к префронтальной коре также развился поздно. Но затем она постепенно начала раскрывать свои тайны великим ученым и таким клиницистам, как Хьюлингс Джексон18 и Александр Лурия19, а в последние десятилетия — Антонио Дамазио20, Хоакин Фюстер21, Патриция Голдман-Ракич22, и Дональд Стас и Фрэнк Бенсон23.
 |
Рис. 4.3. Префронтальная кора |
Командный пункт хорош настолько, насколько хороши его линии коммуникации с боевыми соединениями. Верная своим «управляющим» функциям, префронтальная кора, вероятно, — лучше всех обеспеченная связями часть мозга. Префронтальная кора прямо взаимосвязана с каждой функциональной единицей мозга24. Она соединена с задней ассоциативной корой, высшей инстанцией перцептивной интеграции, а также с премоторной корой, базальными ганглиями и мозжечком, которые вовлечены в различные аспекты моторного управления и движений. Префронтальная кора соединена с дорзомедиальным ядром таламуса, высшей инстанцией нейронной интеграции внутри таламуса, с гиппокампом и относящимися к нему структурами, которые имеют решающее значение для памяти, и с цингулярной корой, которая играет решающую роль для эмоций и для разрешения неопределенности. Вдобавок этот командный пункт соединяется с амигдалой, которая регулирует наиболее базовые отношения между индивидуальными особями, и с гипоталамусом, ответственным за управление жизненными гомеостатическими функциями. Последнее, но далеко не исчерпывающее, — командный пункт связан с ядрами ствола мозга, ответственными за активацию и возбуждение.
Из всех структур мозга только префронтальная кора встроена в такую богатую сеть проводящих путей. Эта уникальная связность делает лобные доли особенно подходящими для координации и интеграции работы всех других мозговых структур — дирижером оркестра. Как мы увидим позднее, эта крайняя степень связности также подвергает лобные доли особому риску повреждения. Как и в политических, экономических и военных организациях, лидер является в итоге ответственным за промахи подчиненных.
Мы увидим позднее, что префронтальная кора, уникальная среди мозговых структур, как бы содержит карту всей коры, — это утверждение сделал впервые Хьюлингс Джексон25 в конце девятнадцатого столетия. Эта особенность префронтальной коры может быть главной предпосылкой сознания, «внутреннего восприятия». Так как каждый аспект нашего внутреннего мира может, в принципе, быть фокусом нашего сознания, разумно предположить, что в мозгу должно существовать место конвергенции всех его нейронных субстратов. Это ведет к интересной гипотезе, что эволюция сознания, высшего выражения развитого мозга, параллельна эволюции префронтальной коры. Действительно, эксперименты показали, что понятие Я, которое является решающим атрибутом сознания, возникает только у высших приматов. И только у высших приматов префронтальная кора занимает ведущее положение в мозге.
| НАЗАД | Оглавление | ВПЕРЁД |